Breastfeeding is an important bonding experience and provides babies with the best start in life. When breastfeeding is not an option, choosing the best infant formula is key for healthy growth and development.
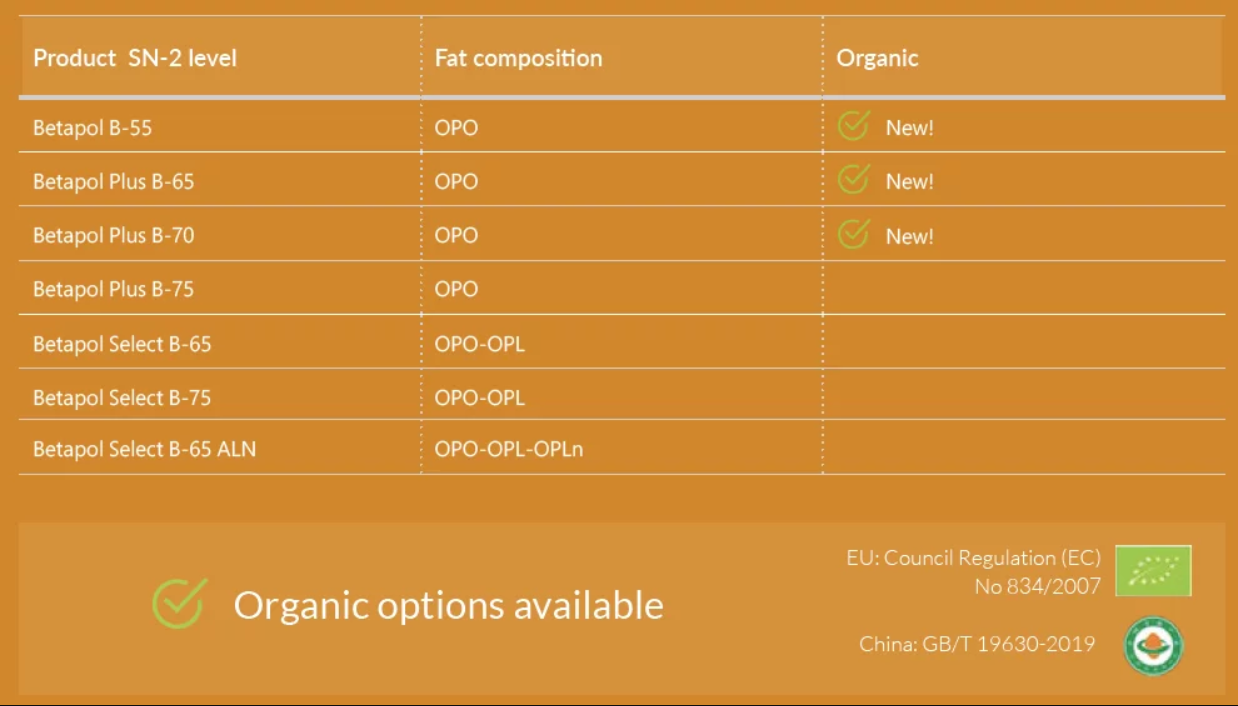
The introduction of Organic Certified Betapol, the first EU Organic and China Organic Standard certified OPO, marks an important milestone.
Grounded in our ambition to be a growth enabler for our customers, Betapol® Organic allows the production of organic infant milk formulas with the clinically proven benefits of OPO.

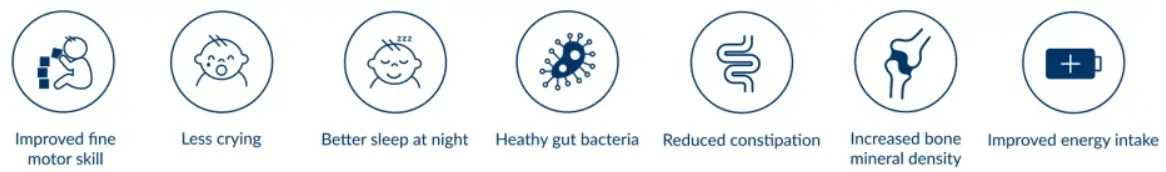

Recent independent clinical research* published in February 2021 indicates a positive effect on the Bifidobacteria level in the infant's gut and fine motor skills. This cluster randomized clinical trial enrolled 199 infants: 66 infants were breast-fed and 133 infants received formula:
- 66 infants received formula containing high sn-2 palmitate with 46.3% of the palmitic acid in the SN-2 position. The content of the SN-2 PA level in the high SN-2 palmitate formula was increased by adding Betapol produced by Bunge
- 67 infants in the control group received formula containing low sn-2 palmitate with 10.3% of the palmitic acid in the SN-2 position from a standard vegetable oil mixture.
The conclusion of the trial states that "Elevating the SN-2 palmitate level in the formula improved infants’ development of fine motor skills, and the beneficial effects of high SN-2 palmitate on infant neurodevelopment was associated with the increased gut Bifidobacteria level."
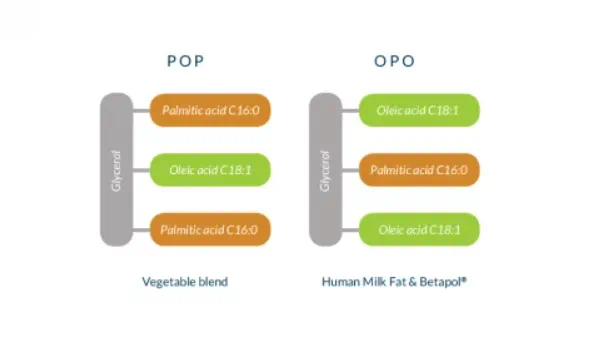
Many standard infant milk formulas have a fat structure (POP) that resembles breast milk fat, but the fatty acids are positioned differently. The fatty acid composition of Betapol mimics the unique OPO structure naturally present in in breast milk. As a result, it optimizes absorption of palmitic acid and calcium which are crucial to energy intake and bone formation.
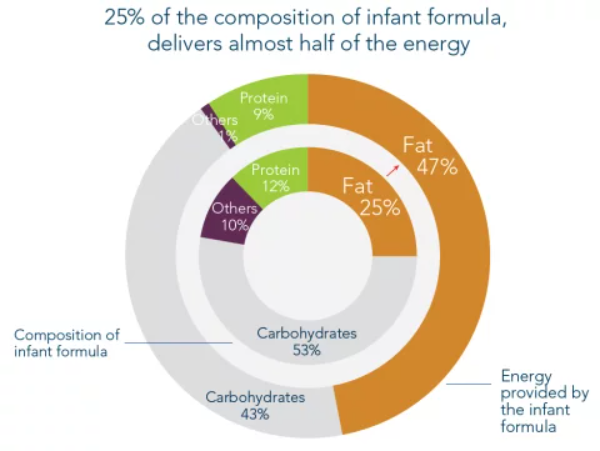
Fats are an essential nutrient for the baby. While mother’s milk fat contains only 3-4% fat it makes up 50% of a baby’s energy intake. Betapol is the ideal choice of fat for infant formula because it mimics the fat composition and unique OPO fat structure that is naturally present in mother’s milk, allowing babies to absorb more palmitic acid and calcium which contributes to significant nutritional benefits. That's why it's widely trusted as the standard for OPO fats.
When we invented OPO, we revolutionized the Infant Formula market by bringing infant formula closer to nature. In 2019 we revolutionized the market again with the introduction of Betapol Plus, the next generation OPO. It is designed to match mothers’ milk fat more closely, by combining the benefits of OPO with the highest OPO/SN-2 palmitate levels available on the market. In 2020 we developed Betapol Select, the first OPL in the market to closely mimic Chinese mother’s milk fat. And in 2021 we are the first company to offer dual Chinese and EU Organic Certification to produce Organic infant formula with Betapol Organic.
*The trial is published on MDPI's website: Nutrients | Free Full-Text | Neurodevelopmental Outcomes and Gut Bifidobacteria in Term Infants Fed an Infant Formula Containing High sn-2 Palmitate: A Cluster Randomized Clinical Trial | HTML (mdpi.com).
Nutra-Ingredients published about the research and its findings here: Infant neurodevelopment: Higher intake of sn-2 palmitate could improve fine motor skills and increase gut Bifidobacteria – China study (nutraingredients.com)
Read our Scientific Summary for a total overview of all clinical studies backing the benefits of OPO.
Connecting farmers to consumers to deliver essential food, feed and fuel to the world.
© 2024 Bunge.